*** Mit Dank an George M. Strain für die fachliche Beratung und die Erlaubnis, Abbildungen aus seinen Veröffentlichungen zu verwenden. ***
Die meisten Fälle von Taubheit beim Hund sind angeboren. Angeborene Taubheit wurde bei mindestens 80 verschiedenen Hunderassen beschrieben. Sie tritt in der Regel gemeinsam mit verschiedenen Formen von Pigmentmängeln auf, die sich in weißen Abzeichen (bis hin zum komplett weißen Haarkleid) oder auch in flächigen Aufhellungen der Grundfarbe äußern können. Weiße Pigmentierung ist unbestritten ein Risikofaktor für Taubheit (Strain GM, 2004) Wir wollen zunächst betrachten, inwiefern Pigmentierung und Hörfähigkeit bzw. deren Verlust bei Pigmentmangel zusammenhängen.
 |
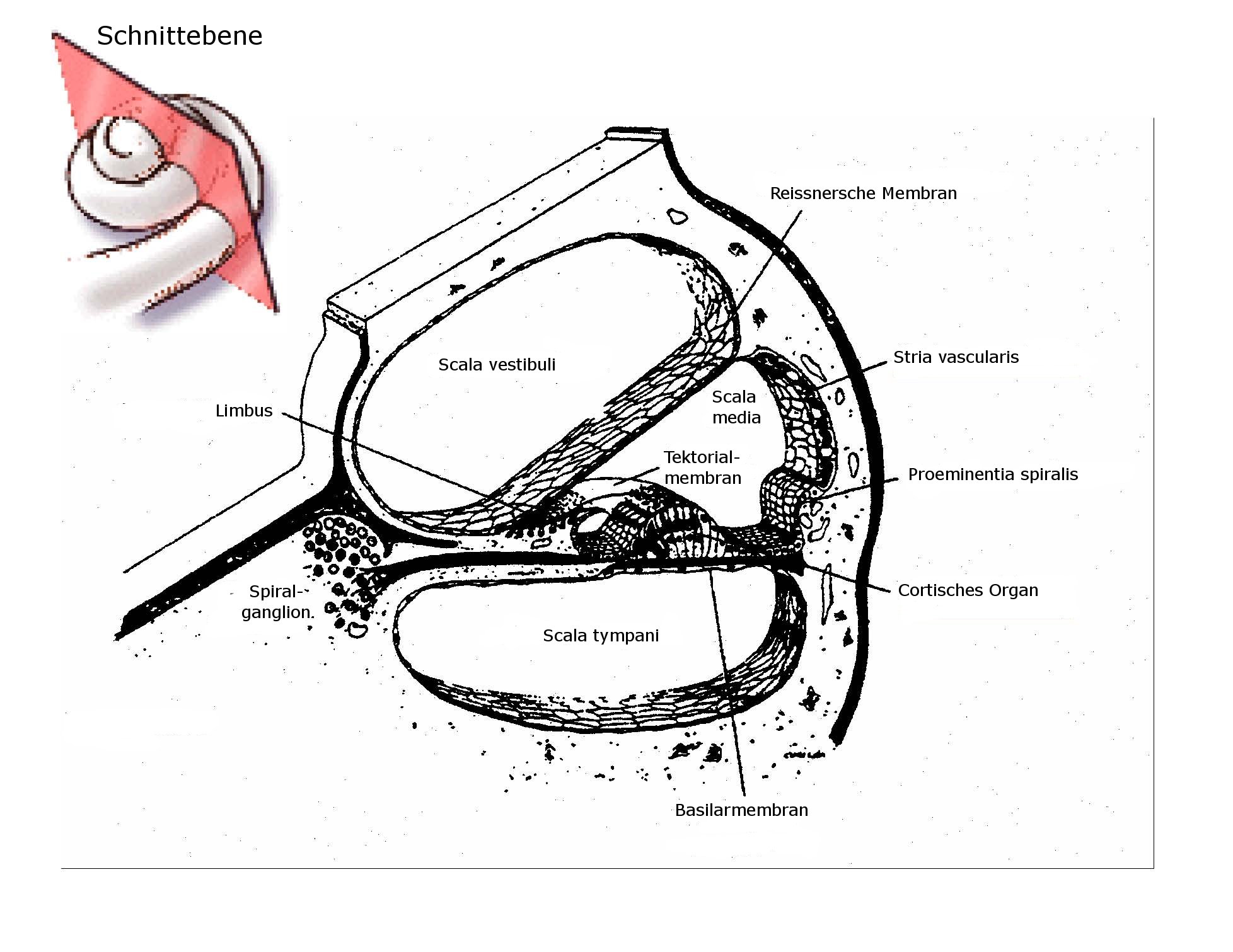
Das Ohr besteht aus drei anatomischen Einheiten: Außenohr, (Ohrmuschel und Gehörgang), Mittelohr (Trommelfell, Gehörknöchelchen und Eustachische Röhre) und Innenohr (Schnecke und Bogengänge). Beim Hörvorgang gelangen Schallwellen über den Gehörgang an das Trommelfell und versetzen dieses in Schwingung. Diese Vibrationen werden über die drei gelenkig verbundenen Gehörknöchelchen Hammer, Amboss und Steigbügel (so benannt aufgrund ihrer prägnanten Formen) an die Schnecke übertragen: Der Fuß des Steigbügels ist über eine Öffnung beweglich mit den drei flüssigkeitsgefüllten Kammern im Inneren der Schnecke verbunden: Der durch die Schallwellen in Vibration versetzte Steigbügel kann so die Schwingungen an die Flüssigkeit in den Kammern übertragen und diese in eine Wellenbewegung versetzen. Diese Übertragung der Schallwellen vom äußeren Ohr bis zur Schnecke des Innenohres funktioniert auch bei pigmentmangelbedingter Taubheit. Der degenerative Vorgang, der zum Verlust des Hörvermögens führt, tritt in der Schnecke selber auf. |
|
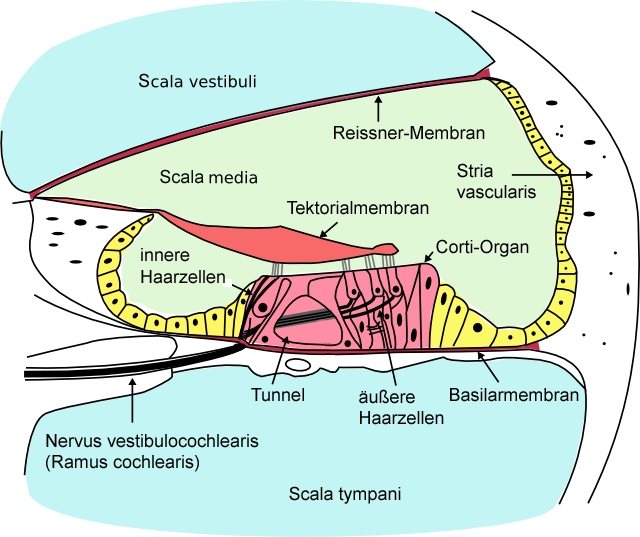
In der Schnecke befinden sich wie gesagt, drei flüssigkeitsgefüllte Kammern (Scala vestibuli, media und tympani im Bild) und die Vibration des Steigbügelfußes versetzt die Flüssigkeit in eine Wellenbewegung. Hier wird die Information über das in der mittleren Kammer (Scala media oder Schneckengang) befindliche sogenannte Cortische Organ mithilfe seiner Sinneszellen, den Haarzellen, weiterverarbeitet: Die Welle wird von den „Haaren“ der äußeren Haarzellen wahrgenommen, die mit der Flüssigkeit in Kontakt sind. Die äußeren Haarzellen übertragen die Schwingungen dann auf die inneren Haarzellen, welche diese wiederum schließlich in Nervenimpulse umwandeln, die zum Gehirn geleitet werden. Damit ist der (sehr vereinfacht dargestellte) Hörvorgang abgeschlossen. Zum genauen Verständnis der Taubheitsproblematik muss noch eine weitere Struktur der mittleren Kammer, des Schneckengangs, erwähnt werden, die sogenannte Stria vascularis: Es handelt sich hier um ein blutgefäßreiches Gewebe an der Außenwand des Schneckengangs (siehe Zeichnung), das an der Produktion der Flüssigkeit beteiligt ist, mit der dieser gefüllt ist. Die Stria vascularis enthält unter anderem auch Pigmentzellen (Melanozyten) …womit wir uns endlich einer Verbindung zum eigentlichen Thema, der pigmentmangelabhängigen Taubheit näheren. |
 |
Die erbliche angeborene Taubheit steht wie gesagt in aller Regel mit der Mutation von Genen in Zusammenhang, die für die Ausbildung von weißen oder aufgehellten Flächen im Fell verantwortlich sind, also dort einen Pigmentmangel hervorrufen.
Die ersten Publikationen zur Taubheit bei Tieren stammen vom Ende des 19. Jahrhunderts und handelten vom Dalmatiner (Rawitz B, 1896) Bis heute ist der Dalmatiner hinsichtlich seiner Hörfähigkeit die am häufigsten wissenschaftlich untersuchte Hunderasse.
|
|
Auf welchem Mechanismus beruht nun die pigmentmangelbedingte Taubheit? Die Ursache liegt in der Degeneration des blutgefäßreichen Gewebes an der Wand des Schneckenganges, der Stria vascularis, die dann auftritt, wenn dieses Gewebe keine Pigmentzellen enthält. In Folge der Degeneration fällt dann die „Decke“ des Schneckenganges (die sogenannte Reissner-Membran, siehe Zeichnung) in sich zusammen und schließlich sterben auch die Sinneszellen des Corti-Organs im Schneckengang ab… also dessen Haarzellen, die, wie oben dargestellt, für das Hörvermögen unverzichtbar sind: Der Hund wird taub, weil die Weiterleitung der Information durch die Zerstörung der Haarzellen an der Stelle der Umwandlung der Schallwellen in Nervenimpulse unterbrochen ist. Es ist bislang nicht bekannt, welche Funktion die Pigmentzellen in diesem Zusammenhang und an diesem Ort genau haben und auch nicht, wieso deren Abwesenheit zur Degeneration des Gewebes führt, aber offensichtlich sind die Melanozyten unverzichtbar zur Funktionserhaltung des Innenohres. Untersuchungen an Dalmatinern haben gezeigt, dass die degenerativen Vorgänge, die zur Taubheit führen, bereits am ersten Tag nach der Geburt der Welpen einsetzen (Johnson LG, 1973). Die durch den Niedergang der Sinneszellen bedingte Taubheit wird jedoch nicht vor der dritten bis vierten Lebenswoche manifest (Strain, 1996). |
Zwei für Variationen der Pigmentierung verantwortlich Gene werden oft mit Taubheit in Zusammenhang gebracht, das Merle-Gen und das Piebald-Gen.
Merle
Beim Merle-Gen sind die Zusammenhänge recht einfach und eindeutig: Während heterozygote Träger des Merle-Alleles zerrissen aussehende Flecken in der Grundfarbe auf einem aufgehellten Grund besitzen, ist der homozygote Merle-Träger überwiegend weiß. Bei Doggen, wo mit dem Merle-Allel ausschließlich im Farbschlag mit schwarzer Grundfarbe gezüchtet wird, werden aus diesem Grunde traditionell die heterozygoten Merle-Träger Grautiger und die homozygoten Merle-Träger Weißtiger genannt. Das Risiko für Taubheit der homozygoten Merle-Träger oder Doppelmerles ist generell erheblich und steigt mit der Vergrösserung des Weißanteil im Fell noch an.
Es soll hier jedoch angemerkt werden, dass auch heterozygote Merle-Träger ein höheres Taubheitsrisiko aufweisen als Hunde, die kein Merle-Allel tragen (Strain GM, 2009). Reetz et al. fanden bei ihren Untersuchungen an 38 Dackeln Störungen des Hörvermögens bei 36,8% der heterozygoten Merle-Träger im Vergleich zu 54,6% bei den Doppelmerle-Dackeln (Reetz I, 1977). Bei der Aufarbeitung der Resultate wurde aufgezeigt, dass diese extrem hohe Anzahl an betroffenen Hunden wohl in erster Linie dadurch zustande kam, dass Reetz et al. auch Hunde mit partiellem Hörverlust dazugezählt hatte, der vermutlich nicht genetisch bedingt war (Strain GM C. L., 2009). In eine aktuelleren Untersuchung an über 2000 Border-Collie-Welpen konnte aber ebenfalls ein signifikant vermehrtes Auftreten von ein- oder beidseitiger Taubheit bei Merle-Trägern nachgewiesen werden: Während nur 6,3% aller untersuchten Welpen Merle-Träger waren, betrug ihr Anteil in der Gruppe der hörgeschädigten Hunde 25%. Bemerkenswert war auch, dass das Risiko für eine taube Mutter, tauben Nachwuchs zu bekommen um das 14fache erhöht war. (Platt S, 2006). In der Untersuchung wurden die Merle-Träger nicht nach homozygoten und heterozygoten Genotypen aufgeschlüsselt. Strain et al. untersuchten 153 Merle-Träger verschiedener Rassen auf deren Hörfähigkeit und Homozyotie bzw. Heterozygotie bezüglich des Merle-Alleles. Er konnte erwartungsgemäß das vermehrte Vorkommen von ein- und beidseitiger Taubheit bei Merle-Trägern bestätigen und insbesondere nachweisen, dass Doppelmerles (25% ein- oder beidseitig taub) signifikant öfter von Taubheit betroffen sind als heterozygote Merle-Träger (3,5% ein- oder beidseitig taub). Strain hebt den erheblichen Unterschied zwischen der Häufigkeit von Hörstörungen bei Doppelmerle-Catahoulas (11,5%) und bei Hunden der übrigen getesteten Rassen (63,6%[!!!]) hervor und führt dies auf die meist starke Pigmentierung der homozygoten Merle-Träger dieser Rasse zurück. Im Weiteren beobachtete Strain ein vermehrtes Vorkommen von Taubheit bei Doppelmerles in Collie-artigen Rassen wie Collies, Shelties oder Border Collies. Inwiefern es hier von Bedeutung ist, dass diese Rassen stets auch Piebald-Träger sind, ist bislang nicht wissenschaftlich untersucht.
Es existiert auch keine wissenschaftliche Publikation zur Häufigkeit von Taubheit bei Grautigern und gefleckten Doggen, beides heterozygote Merle- und zum großen Teil auch Piebald-Träger. Strain benennt jedoch Gefleckte als mit „relativ hohem Risiko für Taubheit, während andere Farbvariationen weniger wahrscheinlich betroffen sind“ (Strain GM, 2004).
Piebald
Piebald gilt als das Gen, welches am häufigsten mit angeborener Taubheit verbunden ist (Strain GM, 2004). Hier ist jedoch anzumerken, dass dies nicht etwa darauf zurückzuführen wäre, dass das Piebald-Gen ein größeres Risiko hinsichtlich angeborener Taubheit darstellt als das Merle-Gen, sondern vielmehr darauf, dass die hochgefährdeten Doppelmerles selten sind aufgrund des in den meisten Rassen geltenden Verbot, zwei Merle-Träger miteinander zu verpaaren. Hingegen gibt es zahlreiche Rassen, bei denen das Piebald-Allel homozygot fixiert ist, in denen also sämtliche Hunde den Genotyp ss besitzen (siehe auch den Artikel zur Vererbung von Piebald).
Bei der Vererbung des Piebald-Gens sind die Verhältnisse komplizierter als beim Merle-Gen : Zwar ist der homozygote Piebald-Träger wie der homozygote Merle-Träger dem größten Risiko für Taubheit ausgesetzt, jedoch gibt es hier erhebliche Rasseunterschiede hinsichtlich der Genwirkung auf die Fellfarbe und des Taubheitsrisikos. Man spricht von variabler Expressivität des Piebald-Gens: Bei verschiedenen Hunderassen, die alle den gleichen Genotyp hinsichtlich Piebald besitzen, gibt es deutlich verschiedene Ausprägungen des Weißanteils im Fell. Unbestritten ist hingegen die Tatsache, dass in zahlreichen Rassen, die piebald-bedingte weiße Abzeichen besitzen, Fälle von angeborener Taubheit stark vermehrt vorkommen. So wurden beim Dalmatiner 29,9%, beim weißen Bullterrier 19,9%, beim Australian Cattle Dog 14,5%, beim Jack Russell Terrier 12,4%, beim Englischen Setter 7,9% und beim „parti coloured“ Englischen Cocker Spaniel 7% der untersuchten Hunde als ein- oder beidseitig taub identifiziert (Strain GM, 2004; Famula TR, 2007).. Bei Rassen, in denen Farbschläge mit und ohne Weißanteil gezüchtet werden, wie Bullterrier oder Englische Cocker Spaniel, ist die Häufigkeit von Taubheit in den Farbschlägen mit Weiß deutlich erhöht (Strain GM, 2004).
Bei Dalmatinern wurde in zahlreichen Studien ein signifikanter Zusammenhang zwischen blauer Augenfarbe und erhöhtem Taubheitsrisiko festgestellt (Holliday TA, 1992; Greilbrokk, 1994; Strain GM K. M., 1992; Famula TR, 2000; Muhle AC, 2002; Strain GM, 2004). Bemerkenswert war hier die Tatsache, dass die Prävalenz in Europäischen Ländern geringer war als in den USA (Muhle AC, 2002; Wood JL, 1997), was möglicherweise mit der Tatsache zusammenhängt, dass in den meisten Europäischen Ländern blaue Augen vom Standard nicht zugelassen sind. Strain et al. wiesen die statistische Signifikanz des Zusammenhangs zwischen blauen Augen und Taubheit außer bei Dalmatinern auch beim Englischen Setter und beim Englischen Cocker Spaniel nach (Strain GM, 2004).
Es ist hier anzumerken, dass Dalmatiner eine spezifische Mutation des Piebald-Gens aufweisen können (zusätzlich zur „typischen“ Mutation aller Piebald-Träger), die bei keiner anderen Rasse nachzuweisen ist (Karlsson EK, 2007). Es ist möglich, dass diese in Zusammenhang steht mit der extremen Häufigkeit der Taubheit bei Dalmatinern (Strain GM C. L., 2009). Aktuelle molekulargenetische Untersuchungen scheinen diesen Verdacht zu erhärten: Dalmatiner, welche die spezifische Mutation aufwiesen, waren signifikant häufiger taub als solche, die sie nicht trugen (Stritzel S, 2009).
Bei Dalmatinern, Englischen Settern und Englischen Cocker Spaniels konnte ein signifikanter Zusammenhang zwischen Hörstatus der Eltern und dem der Nachkommen festgestellt werden. Mit anderen Worten: Hörgeschädigte Eltern hatten hier ein erhöhtes Risiko, hörgeschädigte Nachkommen zu zeugen. Ein besonderes Problem ist dabei die einseitige Taubheit, die ohne einen objektiven Hörtest, den sogenannten BAER (brainstem auditory evoked response)- oder Audiometrie- Test, der die Nervenimpulse in der Schnecke misst, quasi nicht nachweisbar ist. Gelangen aber einseitig taube Hunde in die Zucht, erhöht dies das Risiko für ein- und beidseitig taube Nachkommen und die Anzahl der hörgeschädigten Hunde innerhalb der Rasse steigt weiter an. In gefährdeten Rassen wäre ein solcher Test als unabdingbare Vorbedingung zur Zuchtzulassung empfehlenswert.
Ein Einblick in die Komplexität von möglichen Interaktionen zwischen Farbgenen untereinander und deren Verbindung zur Entstehung von Hörstörungen gibt eine aktuelle Forschungsarbeit zur Taubheit beim Australian Stumpy Tail Cattle Dog, einer Hunderasse, deren Welpen ähnlich wie Dalmatiner weiß geboren werden, und innerhalb einiger Wochen eine rassetypische blaue oder rote Sprenkelung entwickeln, möglicherweise durch das bislang noch hypothetische sogenannte Ticking-Gen hervorgerufen. Vom farblichen Grundtyp her sind diese Hunde stets homozygote Piebald-Träger: Die Sprenklung ist nur auf weißem Grund sichtbar. Es wurde mit 17,8% ein sehr hoher Anteil an ein- oder beidseitig tauben Hunden nachgewiesen. Bei der genetischen Untersuchung ließ sich die ererbte Taubheit einem bestimmten Abschnitt auf dem Chromosom 10 zuordnen. Auf diesem befindet sich ein Gen (dessen molekulargenetische Bezeichnung Sox10 lautet), welches mit dem Piebald-Gen (zur Erinnerung: Molekulargenetische Bezeichnung Mitf) zusammenwirkt und somit in der Entwicklung von Pigmentzellen eine Rolle spielt. Eine Mutation dieses Gens, die mit Taubheit bei den untersuchten Hunden direkt verbunden war, wurde allerdings nicht gefunden. Nichtsdestoweniger gilt das Gen Sox10 als möglicher (aber bei weitem nicht gesicherter) Kandidat, der - vielleicht nur bei gleichzeitigem Vorkommen mit Piebald - angeborene Taubheit verursachen kann. Dies soll nur andeuten, dass auf diesem Feld noch erheblicher Forschungsbedarf besteht.
Mehrere andere Farbgene scheinen nicht mit Taubheit in Verbindung zu stehen. Beim Albinismus beispielsweise sind Pigmentzellen vorhanden, produzieren jedoch kein Pigment. Ein erhöhtes Vorkommen von Taubheit ist bei solchen Hunden nicht festzustellen.
Ungeklärt ist bislang, inwiefern sich das gemeinsame Vorkommen des Piebald- und Merle-Allels (und bei der Deutschen Dogge zusätzlich das Harlekin-Allel) auf das Taubheitsrisiko auswirkt, jedoch ist es laut George M. Strain (persönliche Kommunikation) durchaus wahrscheinlich, dass die Kombination dieser Allele, deren gemeinsame Aktivität in einer Unterdrückung von Pigmentzellen bestehen, die Wahrscheinlichkeit des Auftretens angeborener Taubheit erhöht.
In einem weiteren Artikel wird die ausufernde Verwendung von homozygoten Piebalds in der Doggenzucht kommentiert.
Bibliographie
Famula TR, O. A. (2000). Complex segregation analysis of deafness in Dalmatians. Am J Vet Res., 550-3.
Famula TR, E. J. C. (2007). Heritability and complex segregation analysis of deafness in Jack Russel Terriers. BMC Vet Res., 3:31.
Greilbrokk, T. (1994). Hereditary deafness in the Dalmatian: relationship to eye and coat color. . Journal of the American Animal Association, 170-176.
Holliday TA, N. H. (1992). Unilateral and bilateral brainstem auditory-evoked response abnormalities in 900 Dalmatian dogs. J Vet Intern Med., 166-74.
Johnson LG, H. J. (1973). Vascular anatomy and pathology of the cochlea in Dalmatian dogs. Dans Vascular Disorders and Hearing Defects (pp. 29-95). Baltimore: University Park Press.
Karlsson EK, B. I.-T. (2007). Efficient mapping of mendelian traits in dogs through genome-wide association. Nat Genet., 1321-8.
Muhle AC, J. A. (2002). Further contributions to the genetic aspect of congenital sensorineural deafness in Dalmatians. Vet J., 311-8.
Platt S, F. J. (2006). Prevalence of unilateral and bilateral deafness in border collies and association with phenotype. J Vet Intern Med. , 1355-62.
Rawitz B. (1896). Gehbrorgan und Gehirn eines Weissen Hundes mit blauen Augen. Morphologische Arbeiten, 545-53.
Reetz I, S. M. (1977). [Audiometric findings in dachshunds (merle gene carriers)]. Dtsch Tierarztl Wochenschr., 273-277.
Strain GM. (2004). Deafness prevalence and pigmentation and gender associations in dog breeds at risk. The Veterinary Journal, 23-32.
Strain GM, C. L. (2009). Prevalence of Deafness in Dogs Heterozygous or Homozygous for the Merle Allele. J Vet Intern Med, 282-286.
Strain GM, K. M. (1992). Brainstem auditory-evoked potential assessment of congenital deafness in Dalmatians: associations with phenotypic markers. J Vet Intern Med., 175-82.
Strain, GM. (1996). Aetiology, prevalence and diagnosis of deafness in dogs and cats. Br Vet J, 17-36.
Strain, GM (2011). Deafness in Dogs and Cats, Cabi
Stritzel S, W. A. (2009). A role of the microphthalmia-associated transcription factor in congenital sensorineural deafness and eye pigmentation in Dalmatian dogs. J Anim Breed Genet. , 59-62.
Wood JL, L. K. (1997). Prevalence and prevention of deafness in the Dalmatian--assessing the effect of parental hearing status and gender using ordinary logistic and generalized random litter effect models. Vet J., 121-33.